BrainAGE作为大脑老化的神经影像标志物的十年
随着人口老龄化,神经退行性疾病的发病率越来越高,给个人和整个社会带来越来越大的负担。然而,个体的衰老速度是由环境、基因和表观遗传等各种因素以及各因素间的相互作用决定的。建立神经解剖学衰老过程的生物标志物,是神经科学的一个新趋势,以便在个体水平上,对年龄相关性神经退行性疾病和神经精神疾病进行风险评估和预测。“脑年龄差距估计(Brain Age Gap Estimation,BrainAGE)”方法是基于结构MRI,预测和评估个体脑龄的首个也是实际应用最广泛的概念。本文总结了过去10年内发表的所有研究,这些研究建立并使用BrainAGE方法来评估基因、环境、生活负担、疾病或寿命之间的相互作用,研究衰老对个体神经解剖学的影响。未来,基于结构或功能标记物的BrainAGE和其他脑年龄预测方法可能会改善对神经病学、神经精神病学和神经退行性疾病的个体风险的评估,并有助于开发个性化的神经保护治疗和干预措施。本文发表在Frontiers in Neurology杂志。
引言
随着人口增长和寿命延长,患有一系列(非致命但)致残性疾病(包括认知衰退和痴呆症等神经退行性疾病)的人数正在增加。了解大脑老化过程与神经退行性疾病机制之间的联系,是卫生系统的当务之急,以便制定有效的战略来应对不断增加的负担。衰老被广泛定义为一种时间依赖性的功能衰退,其驱动力是在整个生命过程中细胞损伤的逐渐累积,以及细胞间通讯的变化。衰老也是一个极其复杂的过程,受到多种遗传和环境因素的影响。
为了了解基因、环境、生活方式、健康和寿命之间的相互作用,研究者加速了对个体“生物年龄”的评估,为了:(i)根据预先建立的健康老龄化参考曲线,确定受试者特异性健康特征,以及各种年龄相关疾病的受试者特异性的风险模式,(ii)开发监测(临床)和干预的措施,这些干预措施根据“生物年龄”而不是实际年龄进行个人定制。最近开发了几种基于细胞、组织或功能的生物标记物,用于测量个体衰老过程中的差异,以识别和预测年龄相关疾病的个体风险和死亡率,并改进干预和治疗策略,包括DNA甲基化状态、测量遗传损伤的累积、端粒长度、评估端粒磨损、身体素质以及作为身体、生理和代谢健康衡量指标的非稳态负荷等。
人类大脑结构成熟/老化的特点是区域特异性、非常协调的非线性模式,以及进展和退化/萎缩过程的顺序性,在整个过程中,大脑结构和功能的变化会表现出显著的不同模式,其中一些大脑区域显示出比其他区域更大的变化。随着活体脑成像(特别是磁共振成像(MRI))等非侵入性方法的出现,以及处理和分析MRI数据的复杂计算方法的可用性,关于大脑结构和功能的横截面和纵向神经成像研究,越来越有助于对健康和疾病的大脑结构成熟和老化有更深刻的理解。
随着人们越来越关注衰老与疾病之间的相互作用,越来越多的研究利用神经成像技术来开发个体大脑健康的生物标志物,即所谓的“BrainAGE”。近年来,数据驱动的学习方法,包括交叉验证、模式分类和基于回归的预测分析,是生物医学和神经科学研究的新趋势,这些方法允许研究者在个体水平上进行测量和预测。为了确定大脑成熟和老化的个体轨迹,以及认知功能障碍和年龄相关脑疾病的风险,许多基于大脑结构和功能的年龄或认知状态预测方法在(认知)神经科学中越来越流行,通过识别与预先建立的参考曲线的偏差,或可自动将患有大脑疾病的患者与健康对照组区分开来,来提供大脑结构和功能的个性化生物标记物。这些研究大多使用最先进的机器学习技术在个体水平上进行预测。特别是模式识别和基于回归的计算机建模方法,旨在预测连续变量的值,如结构脑年龄、认知状态或神经心理学特征。这些新的基于大脑的生物标记物,为神经科学在临床实践中的应用提供了一个强有力的策略,并具有广泛的实施范围,例如提供健康大脑成熟/老化的参考曲线,预测个性化的大脑成熟/老化轨迹,发现对大脑健康的保护性和有害性环境影响,从个体大脑结构的疾病特异性变化中分离出与年龄相关的因素,帮助进行风险评估,早期发现某些神经退行性疾病,跟踪个体疾病的进展,以及确定大脑结构老化与认知能力和神经精神症状的个体关系。
“大脑年龄差估计(brain age gap estimation ,BrainAGE)”方法,利用结构性MRI数据直接量化个体大脑老化的加速或减速,是首个评估大脑老化的方法,该方法:
(1)建立了儿童期至青年期健康大脑成熟的参考曲线,以及成年期至老年期健康大脑老化的参考曲线;
(2)检查了神经退行性疾病大脑老化与健康大脑老化参考曲线之间的偏差;
(3)分析了几个样本中个体大脑老化的纵向变化;
(4)使用预测的个体脑年龄与健康脑老化参考曲线的偏差,来预测认知功能的恶化和向阿尔茨海默病(AD)的转化;
(5)研究了若干健康因素和生活方式因素对个体脑老化的影响;
(6)监测保护性干预对个体大脑老化的影响;
(7)也适用于啮齿动物和非人灵长类动物的实验研究。本文首次描述了脑年龄模型的产生,并整合了所有在健康和疾病人群中,使用BrainAGE方法预测个体脑年龄的研究。在可能的情况下,本综述还包括应用其他脑年龄预测方法来探究此问题的研究。表1中列出了本文总结的所有BrainAGE研究的简短摘要。
BrainAGE模型的生成
越来越多的研究正在使用高维神经成像数据,即每个个体通常包括数百个(多模态)参数,并采用监督性的、线性或非线性模式识别技术,以描述和量化整个生命周期中大脑结构的发展和老化。与单变量方法相比,个体大脑结构的多变量分析,能够检测和量化个体年龄的全脑区域或体素结构的细微但广泛的偏差。
一般来说,首先需要训练大脑年龄预测模型,以便随后评估个人的大脑年龄。大脑年龄预测模型是通过识别脑结构年龄和参数的多变量模式,利用大量(认知)健康受试者的MRI数据生成的。随后,将年龄预测模型应用于测试样本,即利用受试者的个体MRI数据估计受试者特定的大脑年龄。个体的预测脑龄与其实际年龄之间的差异,最终确定了个体与典型成熟/衰老轨迹的偏差。
脑龄评估的生成管道
通常BrainAGE模型的工作流包括几个处理步骤(图1)。首先,使用标准化的基于体素的形态计量学(VBM)管道对原始T1加权图像数据进行预处理,从而在以下分析步骤中使用有可比性且更易于处理的数据(参见Preprocessing of raw MRI data )。其次,对预处理的MRI数据进行自动数据缩减,以降低计算成本,避免模式识别的典型方法过度拟合,并提供稳健且广泛适用的年龄估计模型(参见Data Reduction)。第三,进行相关向量回归(RVR),捕捉全脑的多维度的成熟/老化模式,从而构建结构性大脑成熟/老化模型。随后,可以估计个体的脑年龄(参见Training of the BrainAGE algorithm )。
MRI原始数据的预处理(Preprocessing of raw MRI data )
使用SPM对原始MRI数据进行预处理,包括在MATLAB下运行的VBM8/CAT12工具箱。更具体地说,对T1加权图像进行偏置场不均匀性校正。接下来,对图像进行空间归一化。然后,在同一生成模型中将图像分割成脑组织类型,即灰质(GM)、白质(WM)和脑脊液(CSF)。此外,应用自适应最大后验估计和隐马尔可夫随机场模型,来解释部分容积效应。最后,图像预处理包括仿射配准。
数据降维(Data Reduction )
预处理的MRI数据用4或8mm半高全宽(FWHM)高斯核进行平滑处理。此后,数据被重采样到4或8mm的空间分辨率,在删除非大脑区域后,每个受试者分别得到29852或3747个体素。最后,应用主成分分析(PCA)对数据做进一步降维。由于产生的体素中很大一部分仍然与其相邻的体素共享其大部分方差,因此尽管训练样本中的数据集数量低于体素数量,考虑到训练样本中的数据集数量足够,数学上仍允许执行PCA,(参见Performance of the BrainAGE model for brain aging from early into late adulthood)。PCA模型仅在训练数据中计算,随后利用得到的变换参数来降低独立测试样本中的数据维度。
BrainAGE算法的训练(Training of the BrainAGE Algorithm)
BrainAGE框架使用带有线性内核的相关向量回归(RVR)。重要的是,RVR在训练过程中不需要额外的(手动)参数优化,这在计算成本和模型拟合方面优于常用的支持向量机。
一般来说,年龄回归模型是在训练样本中计算的,利用预处理的结构MRI数据作为自变量,实际年龄作为因变量,从而生成健康大脑成熟/老化的复杂模型(图1A,左列)。在该特定回归任务(即,健康大脑成熟/老化)中,计算体素特定的权重,表示该回归任务中的体素特定重要性(关于体素特定权重的图示,请参见图S1中的脑成熟模型和图S2中的脑老化模型)。
随后,应用大脑成熟/老化模型来整合新的受试者全脑中复杂、多维的成熟/老化模式,得出一个单一值,即估计的大脑年龄(图1A,右列)。
最后,预测脑龄和实际脑龄之间的差异,该差异揭示了个体脑龄的差距估计(BrainAGE)得分。对于BrainAGE,正值表示大脑结构成熟/老化提前,而负值表示大脑结构成熟/老化延迟。在纵向研究中,BrainAGE分数的增加表明,随着时间的推移,大脑老化加速。因此,个体BrainAGE评分直接量化了大脑成熟/老化加速或减速的年数(图1B)。例如,如果一个70岁的人的BrainAGE得分为+5岁,那么这个人的典型萎缩模式类似于一个75岁的人的大脑结构。
 图1 对BrainAGE概念的描述。
图1 对BrainAGE概念的描述。
通过VBM对所有的MRI数据自动预处理。(A)使用训练样本的实际年龄以及预处理后的结构MRI数据,训练健康大脑老化模型(左图:年龄回归模型所使用的最重要体素的位置示意图)。随后,根据MRI数据,对额外的受试者的个体大脑年龄进行估计。(B)由预测脑龄和实际年龄之间的差异,得到BrainAGE评分,正BrainAGE评分表示脑老化提前(橙色线),BrainAGE评分增加表示脑老化加速(红线),负BrainAGE评分表示脑老化延迟(绿线)。
常见的BrainAGE模型在参考样本中的交叉验证策略
为了生成和验证大脑年龄模型,大多数研究都采用了所谓的“交叉验证”方法,即使用大部分健康个体参考样本的神经成像参数来生成大脑年龄模型。然后将生成的脑年龄模型应用于模型生成步骤中未包含的参考样本的较小部分(即“left-out,遗漏”),以便根据实际训练样本中确定的神经成像参数来预测个体脑年龄。该程序重复多次,直到为整个参考样本中的每个受试者提供个体脑龄。
为了测量年龄估计的准确性,计算了个体预测脑龄和实际脑龄之间的皮尔逊相关系数(r)、平均绝对误差(MAE)和均方根误差(RMSE):
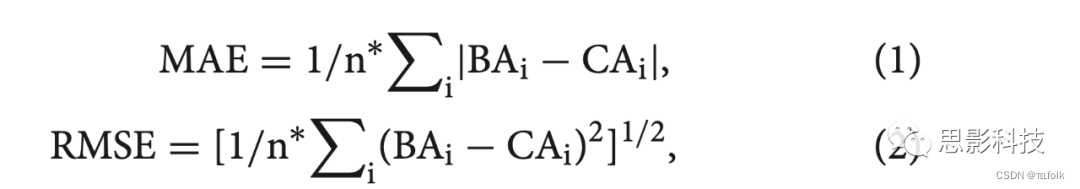
n是测试样本中的受试者数量,BAi是估计的受试者特定的大脑年龄,CAi是受试者特定的实际年龄。此外,使用回归模型的F统计量分析BA和CA之间的拟合。
生成的BrainAGE模型在独立测试样本中的应用
除了参考样本中的交叉验证外,在健康和临床受试者的独立测试样本中进一步验证脑年龄模型,以证明预先建立的脑年龄模型在不同样本甚至不同MRI扫描仪中,具有普适性,这对于脑龄模型在临床环境中的广泛应用,以及在个体水平上诊断和预测脑龄的能力至关重要,被用于监测治疗研究期间大脑年龄的个体变化,或探索各种健康特征、疾病和生活经历对个体大脑衰老的影响。
用于实验动物研究的脑年龄模型的物种特异性适应
狒狒的物种特异性BrainAGE模型
在狒狒的物种特异性BrainAGE模型中,我们使用了Franke等人所描述的定制的预处理管道。为了进一步降低高频噪声,采用空间自适应非局部均值(SANLM)滤波器。分割和空间配准步骤需要一个狒狒特有的组织概率图(TPM)以及一个DARTEL(“Diffeomorphic Anatomical Registration using Exponentiated Lie algebra”)模板,该模板是在一个重新调整的人类模板的迭代过程中估计的。更具体地说,仿射变换最初用于将人类SPM12 TPM和CAT12 Dartel模板映射缩放到狒狒的大脑大小。该模板的图像分辨率设置为0.75 mm的各向同性体素大小。对于执行的每一个迭代步骤,得到的组织映射都是平均的,然后用2 mm FWHM核进行平滑,以估计仿射配准,最后得到一个新的TPM,一个T1平均映射,以及一个狒狒特有的大脑掩模。为了获得平均数据,使用中值函数来减少异常值或处理失败造成的失真。当实际完成的更改低于预定义的阈值(与前一个模板相比)时,迭代过程就会停止,从而产生最终的分割。
分割和配准后,用3mm的FWHM高斯平滑核对数据进行平滑,并重新采样至3mm。最后,应用主成分分析(PCA)进一步降低数据复杂性(如Data Reduction 中所述)。
啮齿动物的物种特异性脑龄模型(Species-Specific BrainAGE Model for Rodents)
如Franke等人所述,用于自动预处理和分析啮齿类动物MRI数据的预处理流程,提供了Paxinos图谱空间的分析,包括几个重新排列和归一化步骤。首先,利用归一化共有信息对Paxinos模板进行仿射共配准。下一步,利用基于变形的形态测量(DBM)方法,来分析实际大脑数据和参考大脑中每个体素之间的位置差异,以检测整个大脑的结构差异。因此,一个动物数据集的所有测量时间点都注册到个体基线扫描。然后,对所有时间点和受试者特定基线测量之间的变形关系进行估计。将基线和后续大脑扫描之间的形态差异最小化,用变形映射编码这些差异的信息。然后,变形的Jacobian(雅克比)行列式可用于计算局部体积变化。最后,用0.4 mm FWHM高斯平滑核滤波每个体素中得到的Jacobian行列式。
技术注释(Technical Notes )
BrainAGE体系是全自动的。所有步骤,包括MRI预处理、数据还原、模型训练和大脑年龄估计,都在MATLAB中执行(www.mathworks.com)。采用SPM8 (www.fil.ion.ucl.ac.uk/spm),集成VBM8工具箱(http://dbm.neuro.uni - jena.de),对T1加权的图像进行预处理。关于狒狒和啮齿动物大脑年龄模型的生成,使用新的CAT12工具箱(http://dbm.neuro.uni- jena.d )。主成分分析应用使用“Matlab降维工具箱”(https://lvdmaaten.github.io/drtoolbox/)。RVR分析是利用“the Spider”工具箱(http://people.kyb.tuebingen.mpg.de/spider/)执行的。
在MAC OS X, Version 10.12, 2.2 GHz Intel Core i7上,每组MRI数据的预处理大约需要20 - 30分钟。训练BrainAGE模型和估计大脑年龄的整个过程总共需要1到5分钟,具体取决于特征、训练和测试对象的数量。
对每个受试者和每次迭代,生成Baboon TPM和模板大约需要30分钟,全部样本(29名对照受试者)总共需要48小时左右。训练狒狒特异性BrainAGE模型,以及估计个体脑年龄的过程总共需要1分钟左右。
在MAC OS X, Version 10.6.3, 2.8 GHz Intel Core 2 Duo上,啮齿动物的MRI数据的预处理时间约为10 - 15分钟,24只大鼠(每只的MRI数据多达13组)预处理时间总共约为5 - 6小时。在这个样本中,训练啮齿动物特异性BrainAGE模型,和估计个体脑年龄的全过程在大约5分钟内完成。
参考样本训练的BrainAGE的预测性能评价
儿童和青少年时期,BrainAGE模型的预测性能
为了生成儿童和青少年时期的脑年龄模型,从儿科MRI数据库(Pediatric MRI Data Repository )[正常大脑发育的NIHMRI研究],中选取394名健康儿童和青少年作为横断面参考样本,年龄5-18岁(平均10.7岁;SD = 3.9年),该数据库中包括从6台不同的MRI扫描仪(1.5T)获得的结构数据。使用“留一”交叉验证,预测脑龄和实际年龄之间的MAE(平均绝对误差)为1.1岁。在预测脑龄和实际年龄之间,87%的方差被解释(r = 0.93;P < 0.001), 95%置信区间在整个年龄范围内是稳定的(±2.6岁;图2 a)。
此外,6个MRI扫描站点中,使用5个站点的数据训练BrainAGE模型,然后应用剩下的一组数据来验证,结果证明在所有的扫描仪位置上,估计精度都保持稳定。预测精度在r = 0.90-0.95和MAE = 1.1-1.3年之间,证明了该脑龄预测模型的稳定性。
到目前为止,已有大量研究建立了大脑成熟模型,年龄范围从幼儿到青年。从参考样本的交叉验证中得出的脑年龄预测,准确性范围为r = 0.43-0.96, MAEs范围为1.0 - 1.9岁。在3-20岁的健康个体的发育过程中,最准确的脑年龄预测模型使用了许多来自不同MRI模式(即T1、T2、DTI)的参数,包括皮层厚度、皮层表面积、皮层下体积、表观扩散系数、分数各向异性、和预先定义的皮质下区域T2信号强度,应用正则多元非线性回归类方法,得到r = 0.96和MAE = 1.0年。尽管每种单一MRI模式在整个年龄范围(即3 - 20岁)显示出相似的预测能力(r≈0.9),但不同模式对大脑年龄模型生成的贡献,在神经解剖结构和年龄范围上有所不同,T2信号强度测量是3-11岁最强的预测因子,扩散系数测量是17-20岁最强的预测因子。此外,与组合模型相比,模式特异性子集的预测精度较差(T1子集:r = 0.91, MAE = 1.7年;T2子集:r = 0.91, MAE = 1.6年;DTI子集:r = 0.90, MAE = 1.7年)。然而,BrainAGE方法仅使用单一MRI模态或单一模态子集就优于所有其他大脑年龄模型,并且额外证明了在不同的扫描仪甚至研究中具有足够的通用性。
BrainAGE模型在成年早期到后期的大脑老化中的表现
在我们首次介绍BrainAGE模型的研究中,我们使用了两种不同的样本来评估大脑年龄,即来自IXI数据库(www.brain-development.org;n = 550,年龄19-86岁,由3台核磁共振扫描仪收集)和另一个独立的健康受试者样本(n = 108,年龄20-59岁,由第4台扫描仪收集)。两个验证样本中健康受试者的大脑年龄被准确估计,因此MAE为5年,总体相关性r = 0.92,预测年龄的95%置信区间在整个年龄范围内是稳定的(图2B)。BrainAGE模型在脑年龄估计上,未显示出系统偏差。此外,大脑年龄估计在性别之间没有差异(两性的r = 0.92;男性MAE = 5.0年,女性MAE = 4.9年)。
另有分析表明,**参考样本中的受试者数量对脑年龄预测的准确性影响最大,即使预处理方法和模型训练算法的选择也会影响模型性能和泛化能力。**具体而言,大脑年龄估计的准确性随着训练/参考样本规模的减少而下降(训练BrainAGE模型的完整数据集[n = 410]: MAE = 5年;1⁄2数据集[n = 205]: MAE = 5.2年;1⁄4数据集[n = 103]: MAE = 5.6年)。结果进一步建议使用仿射配准和相当宽的平滑核,对T1加权MRI图像进行快速的预处理。通过PCA对数据进行降维,适度提高了脑年龄估计的准确性和泛化性,同时加快了生成BrainAGE模型和估计独立被试个体脑年龄值的计算时间(图3)。
到目前为止,已经发表了许多其他的建立大脑老化模型的研究。在健康受试者的整个参考样本中,通过交叉验证得出脑年龄预测,准确性范围为r = 0.43-0.97, MAEs为4.3 - 13.5年,均一误差为5.1 - 21.0年。一般来说,用数学模型模拟健康大脑老化的研究,使用从不同的核磁共振成像模式中获得的许多参数,倾向于提供更准确的大脑年龄预测。在8-85岁的健康参与者样本中,表现最好的模型是基于一系列T1和DTI衍生的参数,利用关联独立成分分析(ICA),总体预测精度为r = 0.97和MAE = 5.9年。另一项研究也使用一系列来自不同的磁共振成像的参数(例如,T1, T2, T2∗,DTI),在20 - 74岁的健康个体的样本中,利用多元线性回归生成和测试大脑年龄模型,整体年龄预测精度r = 0.96。此外,该研究发现体素的平均扩散系数是脑年龄模型的主要预测因子(即,解释62.4%的个体内部方差),其次是GM体积(18.3%)、R2∗ (14.2%)和分数各向异性(3%)。然而,尽管DTI是一种强大的工具,提供了关于组织微观结构和神经纤维连接的独特信息,但从DTI获得的参数会因扫描仪的类型、场强、梯度强度、梯度方向的数量、预处理、拟合程序、牵张算法等而有所差异。然而,包括DTI在内的所有研究都未能在独立的测试样本和扫描仪上证明所建立的脑年龄模型的通用性。
另一项研究使用了来自T1和T2的许多参数,包括皮质和皮质下测量以及连接数据,通过使用线性支持向量回归(SVR),生成和测试脑年龄模型。这种方法在参考样本(组合模型:r = 0.93,MAE= 4.3年)的内部交叉验证中表现很好,但在一个独立的健康受试者的样本(组合模型: r = 0.86, MAE = 8.0年)中进行验证时,不具有普适性。除BrainAGE方法外,在参考样本交叉验证期间以及在独立测试样本中验证脑龄模型期间,利用线性SVR(参考样本:r=0.89,MAE=4.3年;独立测试样本:MAE=3.9年)和高斯过程回归[参考样本:r=0.92,MAE=6.2年;独立测试样本:r=0.93,MAE=5.8年],具有最佳预测准确度。
狒狒BrainAGE模型的性能(Performance of the BrainAGE Model in Baboons )
在建立狒狒特异性脑衰老模型时,仅使用GM(灰质)图像。这个狒狒特有的大脑年龄估计模型,是通过“留一”交叉验证来训练和测试的,每个受试者均使用一次核磁共振扫描。在每个交叉验证循环中,PCA分别在训练集中计算,并在执行RVR之前应用于测试数据。狒狒特异性BrainAGE模型具有很好的准确性(r = 0.80),其中线性回归模型拟合最好(R2 = 0.64;p < 0.0001;图2 c)。MAE为2.1年,与所包括的年龄范围相比,年龄估计误差为11%。
啮齿动物BrainAGE模型的性能(Performance of the BrainAGE Model in Rodents )
正如Franke等人所描述的那样,对啮齿动物特异性BrainAGE模型的训练和测试,采用了受试者特异性留一交叉验证处理。对24只大鼠,在出生后97天至846天之间的13个时间点重复扫描,获得数据集。具体来说,为了模拟啮齿动物特异性的衰老过程,我们对24名受试者中的23名所有扫描时间点的结构MRI数据进行RVR预处理。随后,对被剩下的一只被试每个扫描时间点进行个体大脑年龄的估计,对所有24只大鼠重复整个过程。大脑年龄估计非常准确(r = 0.95;p < 0.0001),线性回归模型显示实际年龄和估计年龄之间的最佳拟合(R2 = 0.91;F = 2622.3;p < 0.0001;图2 d)。平均MAE为49天,与本研究的年龄范围相比,误差为6%。平均RMSE为71天。此外,对大鼠特定的大脑老化轨迹的纵向分析显示,老年大鼠之间的差异在增加(图2E)。
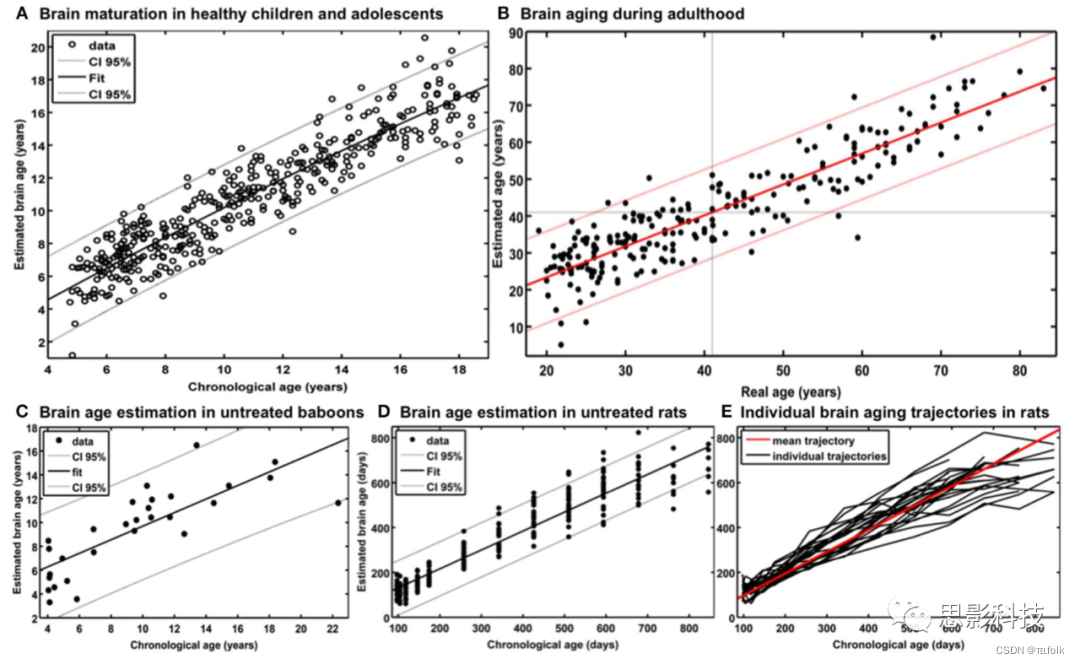
注解:图2 BrainAGE的参考曲线(A) 基于394名健康受试者(年龄5-18岁)的T1图像,预测个体的结构脑年龄。实际年龄显示在x轴上,预测脑龄显示在y轴上。预测脑龄与实际年龄之间的总体相关性为r=0.93(p<0.001),总体MAE=1.1年。二次拟合的95%置信区间在年龄范围(±2.6岁)内是稳定的。(B) 显示整个测试样本的预测脑龄和实际年龄,实际年龄为41岁±11.5岁时的置信区间(红线)。预测脑龄与实际年龄之间的总体相关性为r=0.92(p<0.001),总体MAE=5.0年。(C) 在29只健康对照狒狒的T1 MRI扫描中,根据“留一”交叉验证得出的预测脑龄与实际年龄(以年为单位)的散点图。实际年龄和预测脑龄之间的总体相关性为r=0.80(p<0.001),总体MAE为2.1年。(D)显示了未经治疗的对照组大鼠样本的实际年龄和预测脑龄,包括95%置信区间(灰色线)。实际年龄和预测脑龄之间的总体相关性为r=0.95(p<0.0001)。(E) 大鼠的个体纵向脑老化轨迹。
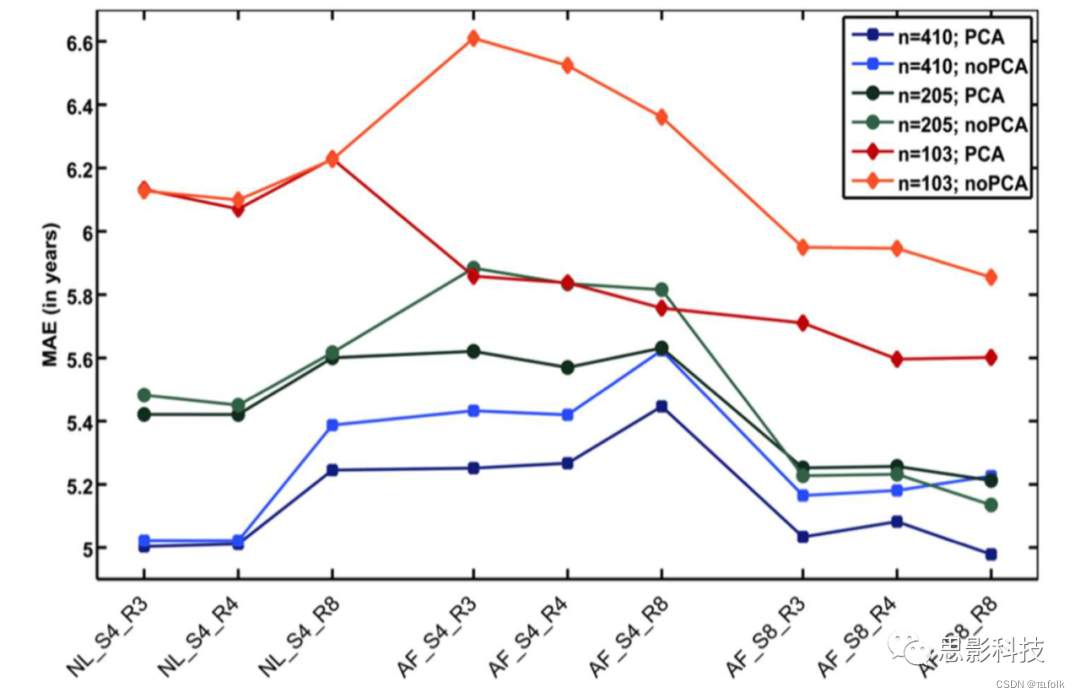
图3 各种参数对BrainAGE估计精度的影响 (1)年龄估计的准确性,主要取决于用于训练年龄评估模型的受试者的数量(蓝线:完整训练样本;绿线:1/2训练样本;红线:1/4训练样本)。(2) T1加权MRI图像的预处理方法对年龄评估的准确性也有很大影响。(3) 通过主成分分析(PCA)进行的数据缩减,仅对平均绝对误差(MAE)产生中度影响。AF,仿射配准;NL,非线性配准;R4/8,重新采样至4/8mm的空间分辨率;S4/8,使用4/8mm的半高宽平滑核进行平滑。
健康成人BrainAGE模型评估的可靠性
BrainAGE评估的扫描—重扫描稳定性相同的扫描仪
为了分析BrainAGE估计的稳定性和可靠性,利用20名健康受试者的T1加权MRI数据,将BrainAGE方法应用于每个受试者的两次MRI扫描,这些扫描在同一台MRI扫描仪(1.5T)上获得,最长时间为90天。结果显示,基于在同一台扫描仪上获得的MRI数据,脑年龄估计具有很强的扫描-再扫描稳定性,第1次和第2次扫描之间的平均BrainAGE分数没有差异(p=0.60),第1次和第2次扫描计算的脑BrainAGE分数之间的类内相关系数(ICC;双向随机单次测量)为0.93[95%置信区间[CI]:0.83–0.97;].
不同MRI场强对BrainAGE评估的影响
为了分析不同扫描仪和场强的模型稳定性,利用60名健康受试者(年龄60–87岁)的T1加权MRI数据,将BrainAGE方法应用于每个受试者的两次MRI扫描(在短时间内通过两台不同的MRI扫描仪(1.5T和3T)获得)。结果表明,场强影响BrainAGE估计,可通过将BrainAGE得分转移到两个数据集中具有线性项的零组平均值,来校正BrainAGE估计,以获得结果的可解释性(图S3)。在对扫描仪特定偏移量进行线性调整后,Student’s t检验未显示1.5T和3T扫描计算出的BrainAGE分数之间的任何差异(p=1.00)。根据1.5T和3T扫描计算的BrainAGE得分之间的ICC结果为0.90(CI:0.84–0.94),表明即使使用来自不同扫描仪和场强的数据,BrainAGE模型仍然具有很强的可靠性和通用性。
BrainAGE对女性的激素相关的短期变化的敏感性
为了建立BrainAGE模型,将其作为一种创新性工具,用来监测并评估由治疗和干预诱导的个体脑老化的短期变化。我们探索了其识别月经周期中由于不同激素影响,而发生的脑结构短期变化的潜力。共有7名年轻、健康、周期稳定的女性(年龄范围21-31岁)在月经期(t1)、排卵期(t2)、黄体中期(t3)和下一次月经期(t4)用1.5T核磁共振进行了扫描。月经周期期间,BrainAGE评分差异显著(p<0.05),从月经期到排卵期,BrainAGE评分下降了−1.3年(SD=1.2年;p<0.05),排卵后缓慢增加(图4)。此外,雌二醇水平与BrainAGE评分呈负相关(r=−0.42,p<0.05),但孕酮水平与BrainAGE无显著相关性(r=0.08,p=0.71)。
Luders等人的一项研究探讨了怀孕后BrainAGE的变化。共有14名健康女性(年龄25-38岁)在分娩后的头两周(产后早期)以及分娩后的4-6周(产后晚期)内接受了3T MRI扫描。BrainAGE评分显著下降,产后早期至晚期平均下降了−5.4年(SD=2.4年;p<0.001)。另外,对激素水平的分析还显示,产后雌二醇(p<0.001)和孕酮(p<0.001)显著降低。
综上所述,这些结果提供了有力的证据,证明月经周期中的激素变化对个体大脑结构有显著影响。BrainAGE方法显示了其捕捉和识别个体大脑结构的细微短期变化的潜力。
BrainAGE模型在儿童和青少年期脑成熟中的应用
早产对个体大脑成熟的影响
在一项针对足月前出生的青少年的研究中,将妊娠27周结束前出生的受试者(即GA(胎龄)<27;n=10)与妊娠29周结束后出生的受试者(即GA>29;n=15)的个体BrainAGE得分进行比较,将预先建立的BrainAGE模型应用于儿童和青少年时期的大脑成熟。在MRI扫描(1.5T)中,受试者年龄在12至16岁之间。结果显示,尽管妊娠年龄的平均差异仅为5周,与出生时GA>29的受试者相比(−0.40±1.50岁),出生时GA(胎龄)<27的青少年组的BrainAGE得分显著降低1.6岁(−1.96±0.68岁),因此这可能意味着大脑结构成熟的延迟。
轻度认知障碍与阿尔茨海默症的BrainAGE
AD患者的脑过早老化
在第一次概念验证应用中,对一组认知健康的对照受试者(CTR;n=232)和一组患有早期阿尔茨海默病的患者(AD;n=102),应用预先建立的成年期BrainAGE模型,进行个体大脑年龄进行的研究。对于AD组,BrainAGE平均得分为+10年(p<0.001),这意味着大脑的系统性提前老化。
在另一项研究中,将预先建立的成年期BrainAGE模型应用于来自Alzheimer’s DiseaseNeuroimaging Initiative (ADNI)数据库的数据,基线BrainAGE评分得出以下组平均值:(1)CTR组− 0.3年(即,在36个月的随访期间,CTR诊断稳定;n=108),(2)sMCI组− 0.5年(即稳定的MCI;在36个月的随访期间,轻度认知障碍(MCI)的诊断稳定;n=36),(3)pMCI组 6.2年(即进行性MCI;在36个月的随访期间,从基线检查时的MC转变为AD;n=112),(4)AD患者6.7年(即在36个月的随访期间或直到死亡,AD诊断稳定;n=150)。事后t检验结果显示,CTR/sMCI组与pMCI/AD组之间存在显著的BrainAGE差异(p<0.05),这强有力的证据表明,pMCI组和AD组的脑结构变化显示了大脑老化提前的模式(图5A)。
CTR、MCI、AD患者个体脑老化的纵向变化
进一步分析探讨了在长达36个月的随访期内,CTR、sMCI、pMCI和AD患者的个体大脑老化轨迹。对于pMCI和AD中的BrainAGE评分,在pMCI中每随访一年(按年龄顺序)大脑老化显著增加1.0年,在AD中每随访一年(按年龄顺序)大脑显著增加1.5年,表明疾病过程中个体脑老化加速(图5C)。由于在基线评估时,pMCI和AD受试者的在基线评估时已提前约6至7年,pMCI和AD的平均随访时间分别为2.6年和1.7年,在最后一次MRI扫描时,两个诊断组的平均BrainAGE评分累积到大约9年(图5B)。相比之下,CTR和sMCI受试者的平均BrainAGE评分在随访期间没有变化,因此表明两组均未偏离健康脑老化。
此外,在36个月的随访期内,结构性脑老化提前与更差的认知功能和更严重的临床症状有关(基线BrainAGE评分:r=0.39–0.46;最后一次随访的BrainAGE评分:r=0.46–0.55)。此外,BrainAGE分数的个体变化与认知测试分数和临床严重程度的个体变化相关(r=0.27–0.33),表明个体脑老化加速与认知功能恶化之间存在显著关系,在pMCI和AD受试者中最为显著。
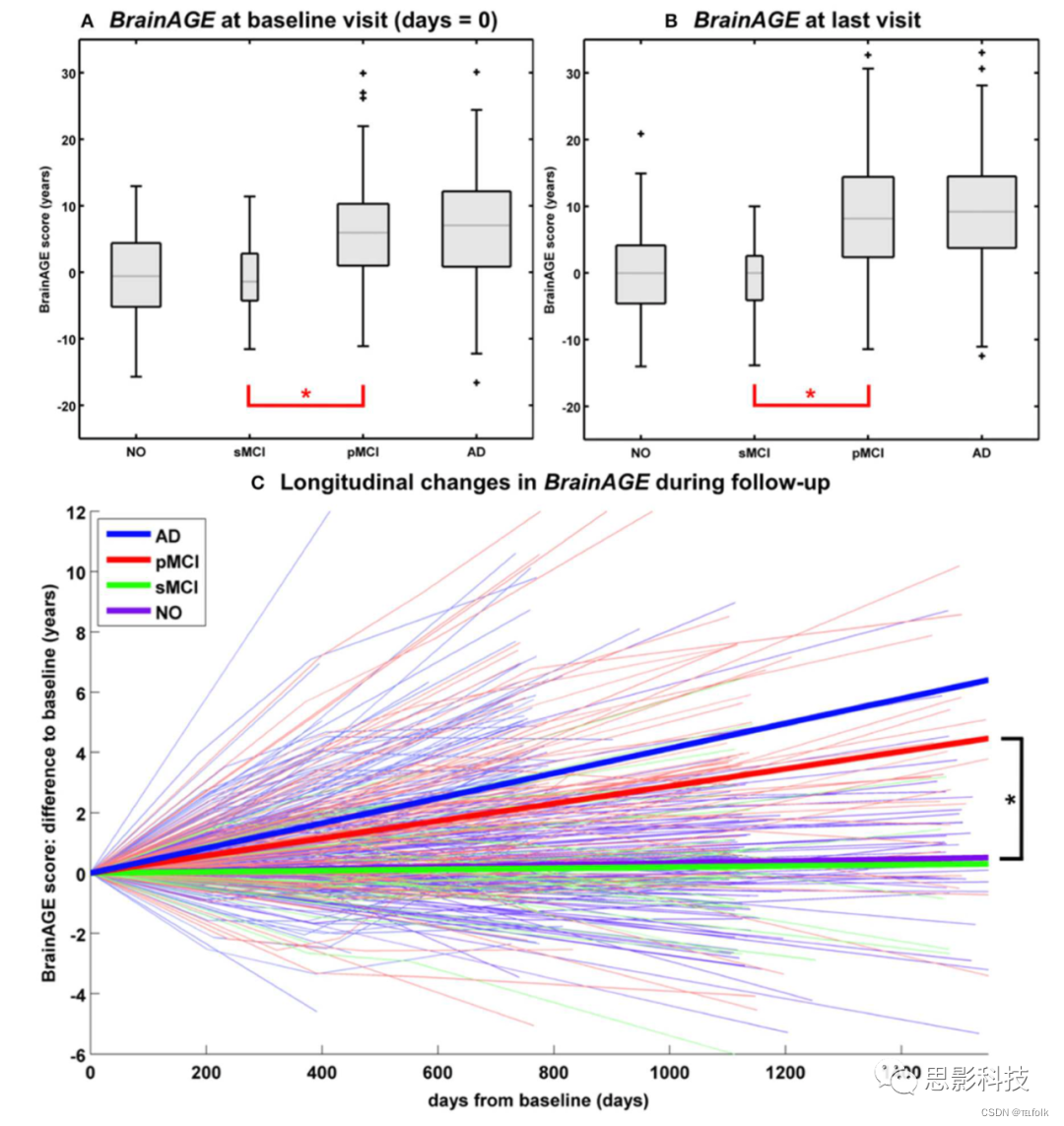
注解:图5 纵向BrainAGE 所有诊断组的(A)基线BrainAGE分数和(B)末次MRI扫描时的BrainAGE分数的箱形图。Post-hoc t检验显示,在两次测量中NO/sMCI与pMCI/AD之间存在显著差异(*p<0.05)。 (C) NO、sMCI、pMCI和AD的BrainAGE分数的纵向变化。细线代表BrainAGE随时间的个体变化;粗线表示各组估计值的平均变化。Post-hoct检验显示,NO/sMCI与pMCI/AD之间的纵向BrainAGE变化存在显著差异(*p<0.05)。
APOE基因型对CTR、MCI、AD纵向变化的影响
在36个月的随访期内,研究载脂蛋白E(APOE)对个体大脑老化轨迹的影响,无论是APOEε4状态,还是特定的等位基因亚型,都对四个诊断组的基线BrainAGE评分没有显著影响。然而,在pMCI和AD组中,APOEε4携带者的个体大脑老化速度明显快于非APOEε4携带者。更具体地说,在pMCIε4携带者中,个体大脑老化加快的速度为每随访一年增加1.1年,而在pMCIε4非携带者中,个体大脑老化加快的速度仅为0.6年左右。同样,在ADε4携带者中,个体大脑老化以每随访一年增加1.7年的速度加速,而在ADε4非携带者中,个体大脑老化仅以每随访一年增加0.9年的速度加速。与之前的结果一致,在健康对照组或sMCI受试者中,无论是ε4携带者还是ε4非携带者,均未观察到与正常脑老化轨迹的偏差(图6)。
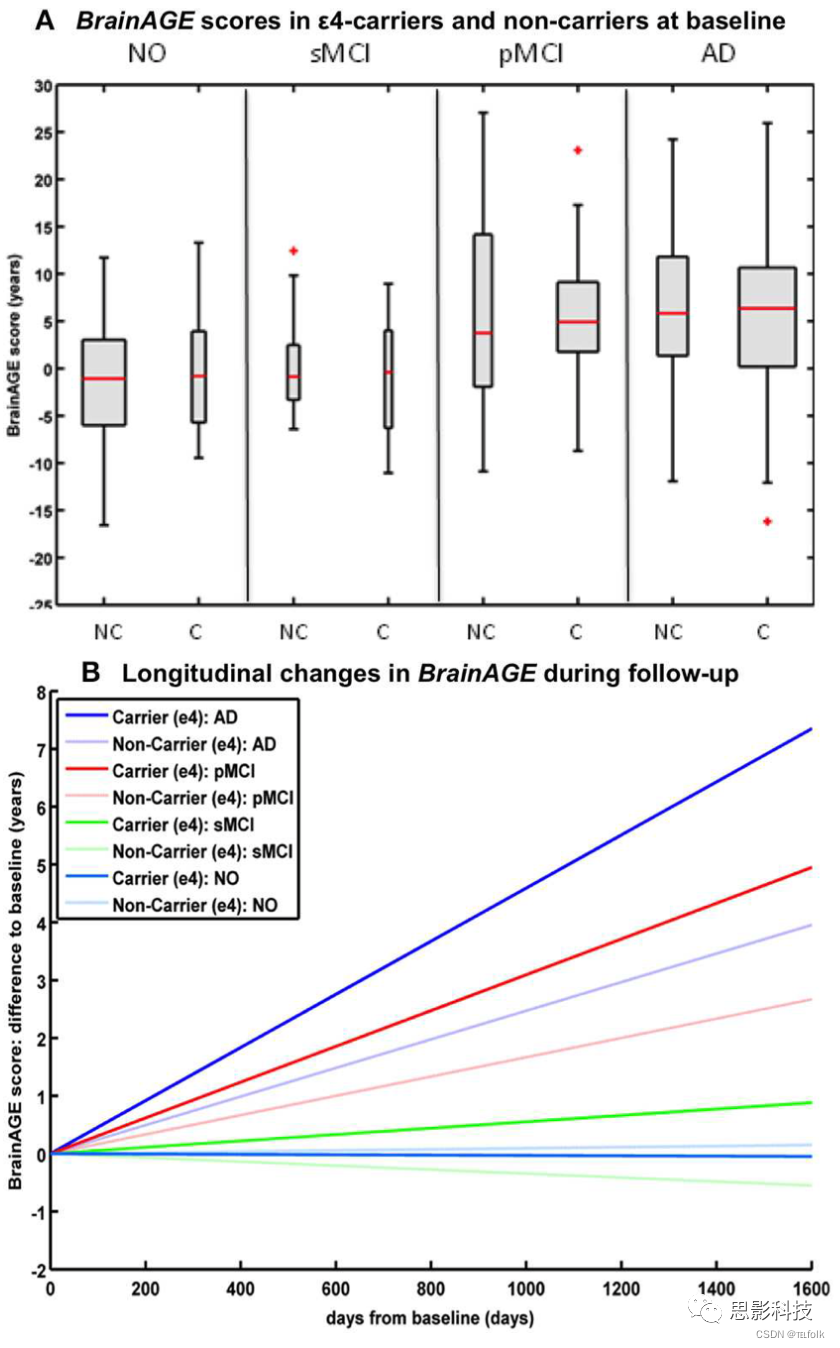
注解:图6 APOEε4-携带者和ε4-非携带者的纵向BrainAGE 在NO、sMCI、pMCI和AD这4个诊断组中,APOEε4携带者[C]和非携带者[NC]在(A)基线时的BrainAGE分数。各诊断组之间,BrainAGE分数存在显著差异(p<0.001)。Post-hoc检验显示,NO和sMCI的BrainAGE得分与pMCI和AD的BrainAGE得分之间存在显著差异(p<0.05)。(B) 4个诊断组(NO(浅蓝色)、sMCI(绿色)、pMCI(红色)和AD(蓝色)BrainAGE得分的纵向变化估计,细分为APOEε4携带者和非携带者。Post-hoc t检验结果显示,ε4携带者和非携带者以及NO/sMCI和pMCI/AD存在显著差异(p<0.05)。
基于BrainAGE预测阿尔茨海默症的转化
基于BrainAGE预测MCI到AD的转化
在Gaser等人的一项研究中,基于结构磁共振成像,用BrainAGE方法提前36个月,预测个体受试者向AD的转化。样本包括195名参与者,基线诊断为MCI,其中133名参与者在36个月的随访期间被诊断为AD。未转变为AD(即sMCI;0.7年)的受试者与在随访的第一年(即pMCI_fast;8.7年)以及随访的第二或第三年(即pMCI_slow;5.6年)内转为AD的受试者相比,其基线检查时的BrainAGE得分之间存在显著差异。研究表明,脑老化提前、认知功能恶化和临床疾病严重程度之间存在密切关系。通过使用基线BrainAGE得分来预测从MCI到AD的转换,后验概率增加到90%。基于基线BrainAGE评分的确定性增加为22%,与基线海马体积(右/左:16%/17%)、认知评分(MMSE:11%;CDR-SB:0%;ADAS:18%)以及最先进的脑脊液生物标记物(T-Tau:4%,P-Tau:0%,Aβ42:0%,Aβ42/P-Tau:8%)相比,该增加是最高的。在随访第一年,基于基线BrainAGE分数,预测未来AD转化率的准确率为81%(受试者操作特征(ROC)分析中的曲线下面积(AUC)=0.83),明显高于基于年龄、海马体积、认知分数和脑脊液生物标记物的转化预测准确性(确切数字见表1)。此外,BrainAGE得分越高,患AD的风险越高,即BrainAGE得分每增加一年,患AD的风险就增加10%(危险率:1.1,p<0.001)。更具体地说,与BrainAGE得分为最低四分位的参与者相比,第二个四分位的参与者患AD的风险大致相同(危险比[HR]:1.1;p=0.68),第三个四分位的参与者患AD的风险高出三倍(HR:3.1;p<0.001),第四个四分位数的参与者患AD的风险高出四倍以上(HR:4.7;p<0.001)(图7A)。BrainAGE优于所有其他基线测量。
APOE基因型对基于BrainAGE预测MCI向AD转化的影响
Loewe等人进一步探讨了在36个月的随访期内,APOE基因型对基于BrainAGE预测MCI转变为AD的影响。与APOE状态无关,基线BrainAGE得分越高,转化为AD的风险越高,若患者BrainAGE得分高于中位数4.5年,则转化为AD的风险比BrainAGE得分低于中位数的患者高出近4倍(HR:3.8,p<0.001)。同样,基于基线BrainAGE得分的Cox回归模型,优于基于认知得分的所有其他模型,即使将APOEε4状态纳入模型(图7b)。此外,基于基线BrainAGE分数的预测,明显比基于实际年龄或认知测试分数的预测更准确(确切数字见表1),尤其是在APOEε4携带者中。
精神疾病对脑老化的影响
最近一项关于精神疾病对个体大脑老化影响的研究,分析了来自精神分裂症(SZ)患者、双相情感障碍(BD)患者(大多有精神病症状或发作)以及21-65岁的CTR参与者的数据。SZ患者的BrainAGE评分显著提高了2.6岁,但BD患者的BrainAGE评分没有明显提高,这表明SZ患者的脑结构发生了提前老化(图8A)。因此,这项研究表明,尽管SZ被称为一种神经发育障碍,但可能存在其他的进行性致病成分。
有趣的是,Hajek等人对患有早期SZ的年轻成人患者、患有早期BD的年轻成人患者和有家族性BD风险的年轻成人(15-35岁)进行的另一项研究得出了类似的结果。特别是,与实际年龄相比,首次发作SZ的受试者的BrainAGE提前了2.6年(p<0.001),而有家族性BD风险或处于BD早期的受试者的BrainAGE与实际年龄没有显著差异,与对照组相比也没有显著差异(p=0.70)。事后分析(post-hoc分析)表明,BrainAGE与全脑的GM总体积呈负相关(图8C)。作者的结论是,BrainAGE有助于BD和SZ的早期鉴别诊断。
对首发SZ的研究调查了并发症肥胖或血脂异常是否会导致大脑改变。与之前的研究相比,首次发作SZ(n=120;18–35岁)的年轻成人参与者表现出神经结构的改变,这导致他们的大脑年龄超过其实际年龄2.6岁(p<0.001)。此外,首发SZ和肥胖的诊断均与BrainAGE相关(p<0.001),首次发作SZ的肥胖参与者BrainAGE得分最高(3.8岁),正常体重对照组最低(−0.3年;图8B)。然而,无论是血脂异常还是药物治疗都与BrainAGE无关。总之,这项研究表明肥胖是弥漫性脑改变的一个独立危险因素,在SZ的早期表现为提前脑老化。因此,针对代谢健康和BMI水平的干预,可能会减缓精神分裂症和精神病患者的大脑老化。
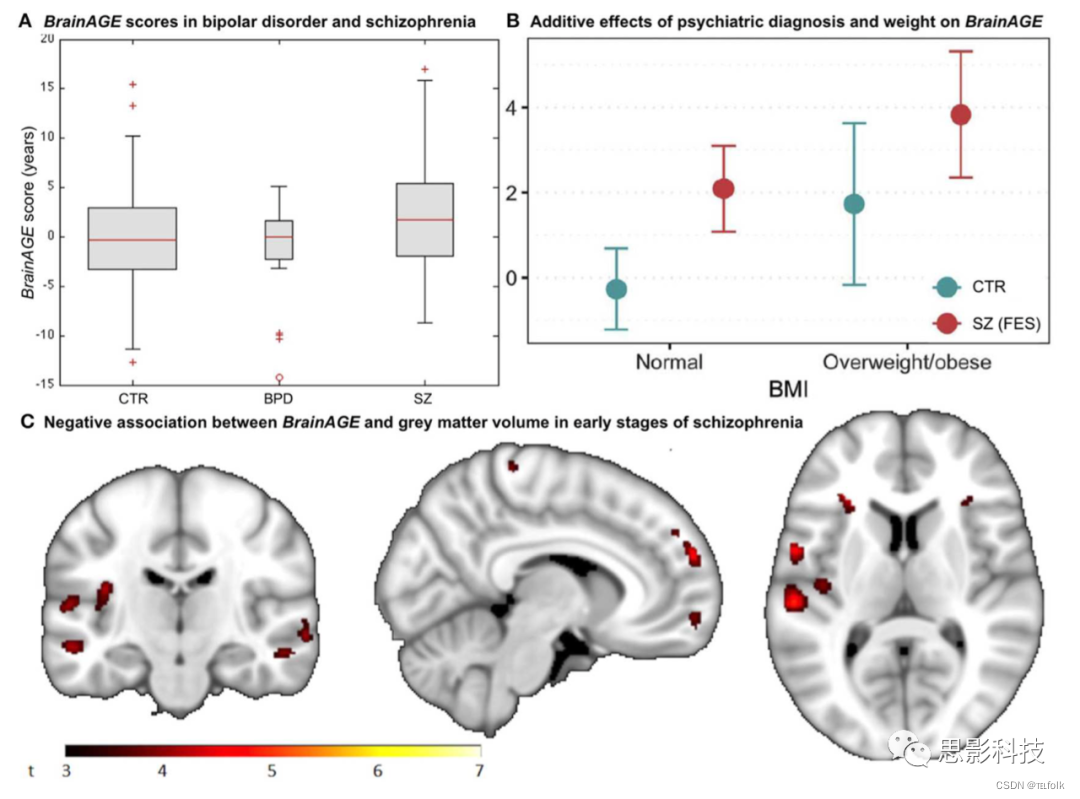
注解:图8 精神疾病中的BrainAGE (A) 健康对照组(CTR)、双相情感障碍患者(BPD)和精神分裂症患者的BrainAGE评分的箱形图,显示显著的群体效应(ANOVA,p=0.009),精神分裂症患者的BrainAGE得分高于CTR或BPD。(B) BrainAGE分数与精神病的诊断以及代谢因素之间的关联。除了首发精神分裂症的影响外,肥胖与BrainAGE分数显著相关。(C) 首发精神分裂症患者的BrainAGE与灰质体积呈负相关(P≤ 0.001)。
个体健康对大脑老化的影响
2型糖尿病对大脑老化的影响
在Franke等人的研究中,将BrainAGE方法应用于2型糖尿病(DM2)参与者和CTR参与者(平均年龄:65±8岁)的样本,以量化DM2对认知健康老年人的大脑老化的影响。与年龄匹配的健康CTRs(健康对照)相比,DM2参与者的BrainAGE显著增加了4.6年(p<0.001)。此外,糖尿病持续时间越长,BrainAGE得分越高(r=0.31,p<0.05)。此外,BrainAGE评分也与空腹血糖呈正相关(r=0.34,p<0.05),最低值与最高值的参与者之间的差异为5.5年(p<0.05)。
2型糖尿病对大脑老化的纵向影响
此外,Franke等人进一步分析了DM2和CTR参与者中的一小部分样本,这些参与者在基线评估后3.8±1.5年完成了后续MRI扫描。两组之间或两个时间点之间的GM和WM体积没有差异。然而,在DM2患者中,BrainAGE评分每随访一年增加0.2年,但在随访期间CTR没有变化。具体而言,与CTR相比,DM2患者的基线BrainAGE得分增加了5.1年(p<0.05),在随访期间,他们甚至增加了0.8年(p<0.05)。因此,在随访期间,DM2中的大脑老化速度甚至更快。
个体健康与大脑老化
除了DM2对非痴呆老年人大脑老化的影响外,Franke等人的研究还探讨了生活方式风险因素(即吸烟时间、饮酒量)、个体健康标志物(即高血压、TNFα)以及常见临床结果(即认知、抑郁)的影响。结果显示,BrainAGE与吸烟时间(r=0.20,p<0.01)、饮酒量(r=0.24,p<0.001)、TNFα水平(r=0.29,p<0.01)、语言流利度(r=−0.25,p<0.01),抑郁(r=0.23,p<0.05)相关,但与高血压无关(p=0.9)。此外,将这些测量值中最低值(即第1个四分位数)的个体与最高值(即第4个四分位数)的个体进行对比,对于吸烟时间,BrainAGE差异为3.4年(p<0.01),对于饮酒时间,BrainAGE差异为4.1年(p<0.01),对于TNFα,BrainAGE差异为5.4年(p<0.01),对于语言流利程度,BrainAGE差异为5.6年(p<0.001),对于抑郁,BrainAGE差异为5.4年(p<0.01;图9A),所有结果与糖尿病病程、性别和年龄无关。
健康特征对大脑老化的性别特异性影响
Franke等人的一项研究利用60-90岁认知未受损参与者的样本,进一步探讨和量化了个体健康的各种生理和临床标志物对个体BrianAGE分数的影响。
在男性样本中,纳入的健康参数解释了39%的BrainAGE差异(p<0.001),其中体重指数(BMI)、尿酸、γ-谷氨酰转移酶(GGT)和舒张压(DBP)贡献最大。另外的四分位分析显示,第1和第4个四分位组之间的BrainAGE得分存在显著差异(图9B,左图),BMI为7.5年(p<0.001),DBP为6.6年(p<0.01),GGT为7.5年(p<0.01),尿酸为5.6年(p<0.05)。当结合这四种健康指标时,对个体BrainAGE的影响甚至是复合的。详细地说,比较低于BMI、DBP、GGT和尿酸中位数的男性受试者和高于BMI、DBP、GGT和尿酸中位数的男性受试者的个体脑年龄,得出的BrainAGE得分分别为−8.0岁和6.7岁(p<0.05;图9B,右图),因此表明男性个体健康与神经结构老化之间存在密切关系。
在女性样本中,纳入的健康参数解释了32%的BrainAGE差异(p<0.01),其中GGT、天门冬氨酸氨基转移酶(AST)、丙氨酸氨基转移酶(ALT)和维生素B12贡献最大。此外,第1和第4个四分位组之间的BrainAGE得分存在显著差异(图9C,左图),GGT为6.6年(p<0.01),AST为3.1年(p<0.10),ALT为5.1年(p<0.05),维生素B12为4.8年(p<0.05)。同样,当结合这四个健康标志物时,对个体BrainAGE的影响是复合的,平均BrainAGE得分为−1.0岁与3.8岁(p<0.05;图9C,右图),因此表明女性个体健康与神经结构老化之间的关系与男性类似。
大脑老化的保护干预
长期冥想练习对大脑衰老的影响
为了探索长期冥想练习的效果,Luders等人的研究包括50名冥想练习者(有4-46年的冥想经验,平均值:20±11年)和50名非冥想、年龄匹配的CTRs。在50岁时,冥想练习者的BrainAGE比CTRs低约7.5年(p<0.05)。此外,性别也发挥了主要作用,女性的BrainAGE比男性低3.4岁(p<0.01)。此外,各组之间的年龄交互作用是显著的(p<0.05),后续分析显示冥想练习对BeainAGE有显著影响。具体地说,在50岁以上的年龄段中,每年冥想练习者的BrainAGE显著减少1个月22天(图10)。
音乐制作对大脑老化的影响
另一项研究调查了音乐制作对大脑老化的影响,包括25±4岁的非音乐家、业余音乐家和专业音乐家。这三组人在年龄、性别、教育程度和其他休闲活动方面都非常匹配。“音乐家身份”对BrainAGE有显著影响(p<0.05;非音乐家:−0.5±6.8年;业余音乐家:−4.5±5.6年;专业音乐家:−3.7±6.6岁),表明音乐对个体大脑老化的减速作用。事后(post-hoc)比较显示,与非音乐家相比,业余音乐家(p<0.05)和专业音乐家(p=0.07)的BrainAGE得分较低。而在业余音乐家中,音乐活动年限与BrainAGE得分之间没有显著相关性(r=−0.1,n.s.),在专业音乐家中发现了很小的相关性(r=0.3,p<0.05)。因此,音乐制作似乎对大脑老化有减缓作用,特别是对业余音乐家而言,而专业音乐家则显示出较低的影响,可能是由于压力相关因素的干扰。
产前营养不良对个体脑老化的性别特异性影响
人类研究的结果
最近的一项研究利用Dutch famine birth cohort 的子样本,调查了妊娠早期胎儿营养不良对晚年个体大脑老化的影响。MRI子样本中的参与者在采集MRI图像时年龄约为67岁,包括1944/45年冬季饥荒前出生的个体、妊娠早期暴露于饥荒的个体以及饥荒后受孕的个体。在女性中,67岁时观察到的BrainAGE差异中,28%是由出生特征、MRI数据采集时的实际年龄和饥饿暴露解释的(p<0.05),而在男性中,76%的BrainAGE差异是由出生特征、晚年健康特征、实际年龄、饥饿暴露解释的(p<0.05)。在男性中,各组之间的BrainAGE得分存在显著差异(p<0.05)。在女性样本中,各组之间的BrainAGE得分没有差异。在男性中,事后检验(post-hoc test)显示,怀孕早期暴露于饥荒的个体的大脑老化提前了2.5年(p<0.05),而那些在饥荒之前出生的人的大脑老化延迟了2.5年−1.8年,结果相差约4年(p<0.05;图11A)。关于BrainAGE得分,男性和女性之间没有显著差异。
非人类灵长类动物的研究结果
一项关于狒狒母体营养限制(MNR)的实验研究,也基于狒狒特有的BrainAgE模型,研究了产前营养不足对结构性脑老化的影响[见Species-specific BrainAGE model for baboons ]。实验组包括11名受试者[5名雌性],在整个妊娠期间,MNR导致的产前营养不良率为30%。CTR组包括12名年龄相同的受试者[5名磁性]。在MRI数据采集时,受试者的年龄为4-7岁[人类相当于14-24岁]。在雌性MNR后代中,与雌性CTR后代相比,狒狒特定的BrainAGE分数增加了2.7年(p=0.01;图11B),这表明在整个妊娠期,由于产前营养不良导致大脑过早老化。MNR和CTR雄性后代的BrainAGE得分没有差异。
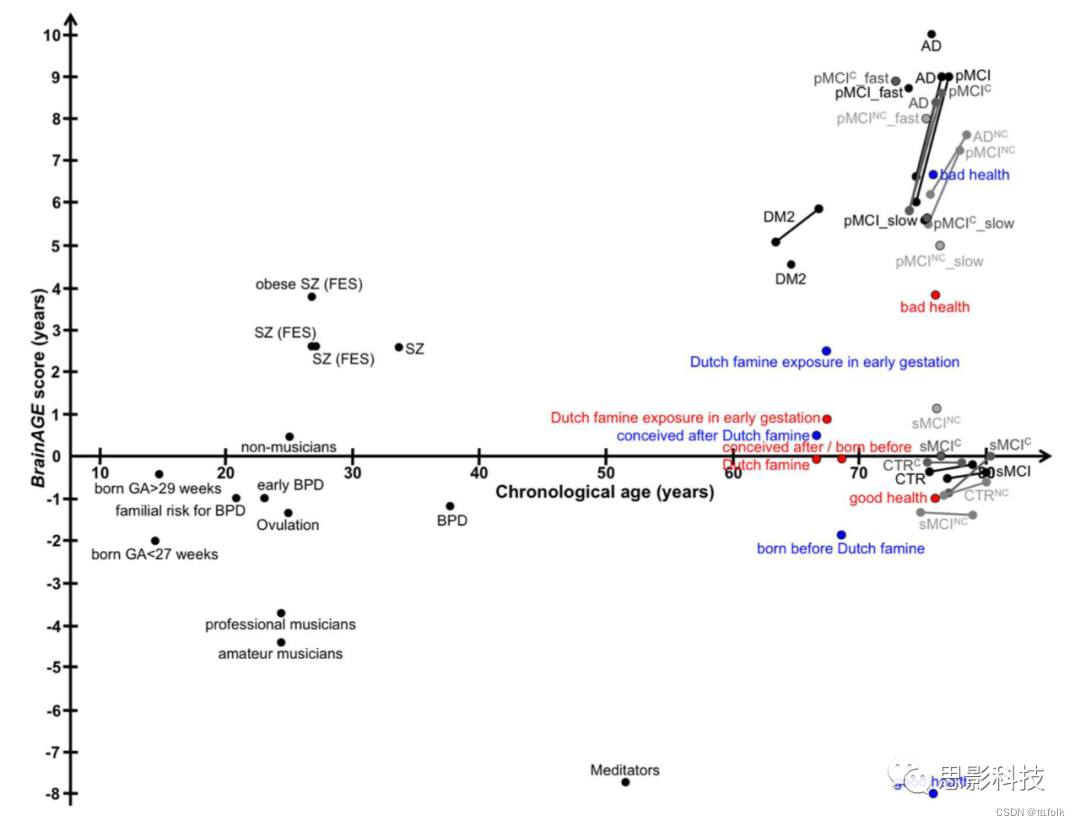
注解:图12 人类研究中BrainAGE结果的图形摘要点,研究手段;直线、纵向结果;蓝色,男性;红色,女性。
【AD,阿尔茨海默病;BPD,双相情感障碍;CTR,对照受试者;DM2,2型糖尿病;FES,精神分裂症谱系障碍的首次发作;GA,胎龄;MCI,轻度认知障碍;pMCI,进行性MCI(即在随访期间从MCI转换为AD);
pMCI_快速,基线诊断为MCI,在前12个月内转为AD;pMCI_缓慢,基线诊断为MCI,在前12个月的随访后报告转为AD;sMCI,稳定的MCI(即在所有可用时间点诊断为MCI,但至少持续36个月);SZ,精神分裂症]。
总结:
本文回顾了利用创新的BrainAGE生物标记物捕捉个体年龄相关的大脑结构的研究,涵盖了从童年到老年的年龄范围(图12为人类研究所有结果的图形摘要)。这种预测性分析方法提供了一种个性化的大脑结构生物标志物,有助于阐明和进一步研究大脑结构和疾病状态中个体差异的模式和机制。因为脑年龄估计是在个体水平上进行的,脑年龄生物标记物可能非常适合临床应用。该方法是从多元模式和整个大脑体素之间的相互作用中得出个体预测。与区域或整体体积、皮质厚度或部分各向异性等其他结构测量相比,BrainAGE分数保留了大脑结构及其区域相互作用中细微变化的复杂模式。此外,将来自整个大脑的复杂多元结构信息简化为一个单一指标,解决了多重比较的问题,并能够更好地检测效果。
根据美国老龄研究联合会(the American Federation of Aging Research ),老龄化标志物应具有某些特征:它们应能够确定生物学老化,预测老化速度,监测老化的基本过程,并准确、高效、重复地测量,而不会对受试者造成伤害。此外,标记需要适用于整个物种的机械检查。然而,由于提取方法、实验室相关方法的细节和测量方法的差异,一些广泛使用的衰老生物标志物(如端粒长度)的重复性和准确性差异很大。因此,准确性有时很低,以至于测量误差阻碍了端粒长度差异的检测。尽管衰老的生物标志物最好与机械性衰老过程密切相关,但与大脑功能和结构相关的大脑衰老标志物的开发更为先进,并与年龄和诊断特异性有相当高的相关性。此外,基于结构MRI的大脑老化标记物显示,实验室或研究地点的个体间差异和测量方法差异较小。表型相关标记物的优越性可以由以下几个原因来解释:目前,由于大脑老化的过程复杂且尚未完全理解,因此更容易确定表型。生物环境中的许多代偿途径更是如此,生物体通过这些代偿途径调节或响应衰老过程。除了存在于细胞水平的复杂性外,生物体可以对无限多的生物和环境影响作出反应,而表型的变化有限。因此,建立脑结构成熟和老化(如脑老化)的表型相关生物标记物,可能是评估和纵向跟踪个体脑老化轨迹的更好方法。
一般来说,认知障碍不仅仅是由一种疾病引起的。认知障碍可能由AD和其他形式的痴呆症以及几种疾病,如创伤性脑损伤、中风、抑郁症或发育障碍引起的。随着心理健康被视为限制老年人生活质量的主要决定因素,与年龄相关的认知能力下降在现代社会中日益受到关注。因此,测量个体大脑年龄和预测个体认知衰退轨迹的生物标记物是非常可取的。基于结构神经成像数据确定大脑年龄的方法,旨在通过建立健康大脑年龄的可靠参考曲线和提供个体大脑年龄测量,同时考虑大脑中的多维度萎缩模式,指示大脑结构中年龄相关变化的偏差。尽管多种因素影响和改变个体的大脑老化轨迹,但正常的大脑老化遵循GM和WM丢失以及CSF扩展的协调和顺序模式。一些研究基于MRI的结构性脑老化模型,证明脑过早老化与AD疾病严重程度和认知功能下降、MCI和AD、向AD转化、SZ、创伤性脑损伤、HIV、慢性疼痛、DM2、孕期营养不良,以及较差的身体和精神健康、非稳态负荷较高以及死亡率增加的迹象之间存在着深刻的关系。此外,个体大脑老化与各种健康参数、个人生活方式或药物使用、教育和体育活动水平以及冥想练习之间存在显著关联。然而,尽管Brown等人的研究表明,青少年大脑早熟程度的增加与执行智能测量的增加之间存在关系,Steffener等人的研究也表明,成人大脑老化延迟与高等教育水平之间存在相关性,这一问题必须在认知储备和IQ水平方面,通过具有良好特征和经过良好测试的样本进行更深入的探讨。
总之,这里介绍的表型方法已经建立并验证了与年龄相关的大脑结构变化的参考曲线。此外,它还显示出在多中心研究中易于应用的巨大潜力。因此,这种预测性分析方法为确定大脑结构的生物年龄提供了个体化的生物标志物,这也与认知功能有关。这种基于MRI的标记物能够预测大脑成熟和老化的个体差异,以及与年龄相关的认知能力下降和神经退行性疾病的发生。本文阐述了神经影像学数据可用于建立脑老化生物标志物的证据,这种生物标志物被证实可提供重要的预后信息。未来,结合结构和功能性脑年龄的不同生物标志物,如相比于实际年龄,在各种神经和精神疾病以及神经退行性疾病中,基于“视觉注意理论”的年龄相关变化的参数评估,可提高检测生物年龄差异的敏感性和特异性。结构和功能性脑年龄评估中包含的重要预后信息,可能有助于制定个性化的神经保护治疗和干预措施。